



2020年3月17日公開
担当:上野将紀 先生
所属:システム脳病態学分野
私たちは普段,何気なく動作しているが,この動作が,脳,もっと言えば,神経細胞でつながった回路によって制御されているというのは,よくよく考えてみれば驚きである.脳は外部環境からの様々な五感情報の入力,あるいは自分自身に湧きおこる内的な動機を認知し,それに適応した出力を導きだす精巧な臓器といえる.出力にあたる主な要素が運動で,突きつめていえば,筋肉を動かすことにある.運動は多様だ.喉の渇きを感じコップへ手を伸ばし,熱いものを触れば手を引っ込める.職場に向かって歩き,ピペットを操作する.話し笑うことも,バスに揺られ座っている姿勢や呼吸すらも筋肉の所作といえる.この間に行われる運動の選択がどのようになされ,また600を超えるともされる全身の筋肉をどうやって協調的に動かせるのか.運動とその神経の仕組みはきっと精密で複雑であろうが,それゆえ謎が多く,長い歴史をもって人々を惹きつけてきた.一方,脳研究所の使命である脳の病気克服の観点からは,脳卒中や脊髄損傷,ALS,パーキンソン病など,運動の機能を侵し日常生活を困難にする多くの病気の存在がある.しかしいずれも根本的な治療法は見出されていない.病態の異常と健常時の正常は,裏返しの関係である.こうした疾患を克服するには,病態の原因を探ることが重要で,一方,運動がなされる神経の機構の理解も不可欠である.
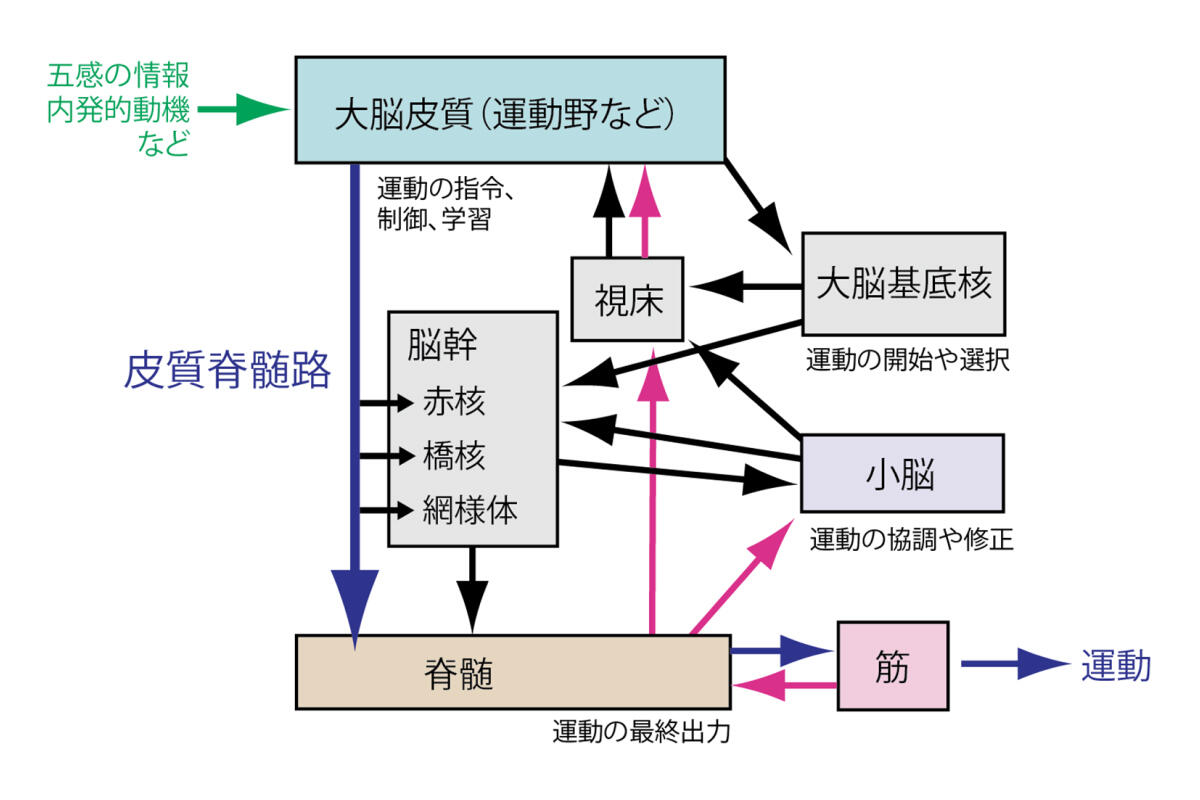
図1.運動回路の概要
運動は,多様な脳領域に担われる(図1)1.最上位に位置する大脳皮質は,運動の制御と指令,学習の中枢で,その指令は直接,あるいは脳幹部を介して下行し脊髄へ伝わる.脊髄には,筋肉へつながる運動ニューロンがあり,筋肉を収縮させる最終出力の地点となる.運動はさらに,小脳,大脳基底核,という2大システムと関わり,適切な動作が生み出される.小脳は,他の脳脊髄領域から体の位置など運動中の情報を受け取り,その修正と学習によってスムーズな動作に貢献する.大脳基底核は,運動の開始や停止を適切に選択し動作をスムーズにする.この大きな脳内回路の枠組みに,運動を生み出すメカニズムが備わっている.しかし,個々の領域内にある数百億から数千億ともいわれる神経細胞の数を考えてみると,それらが一体どのようなつながりと活動によって,無数の動作パターンを生み出すのか,結局のところよくわかっていない.
わたしたちは,この中でも特に,大脳皮質と脊髄を直接結び,運動を制御する「皮質脊髄路」に興味をもち,研究を進めてきた.この回路は,最上位中枢から最下位へ指令をだすシンプルな運動経路ともとらえられるが,実際は複雑で,多様な動作に関わる正確な仕組みは今もってわからないことだらけである.その研究の歴史と課題を,本コラムで紐解きたい.
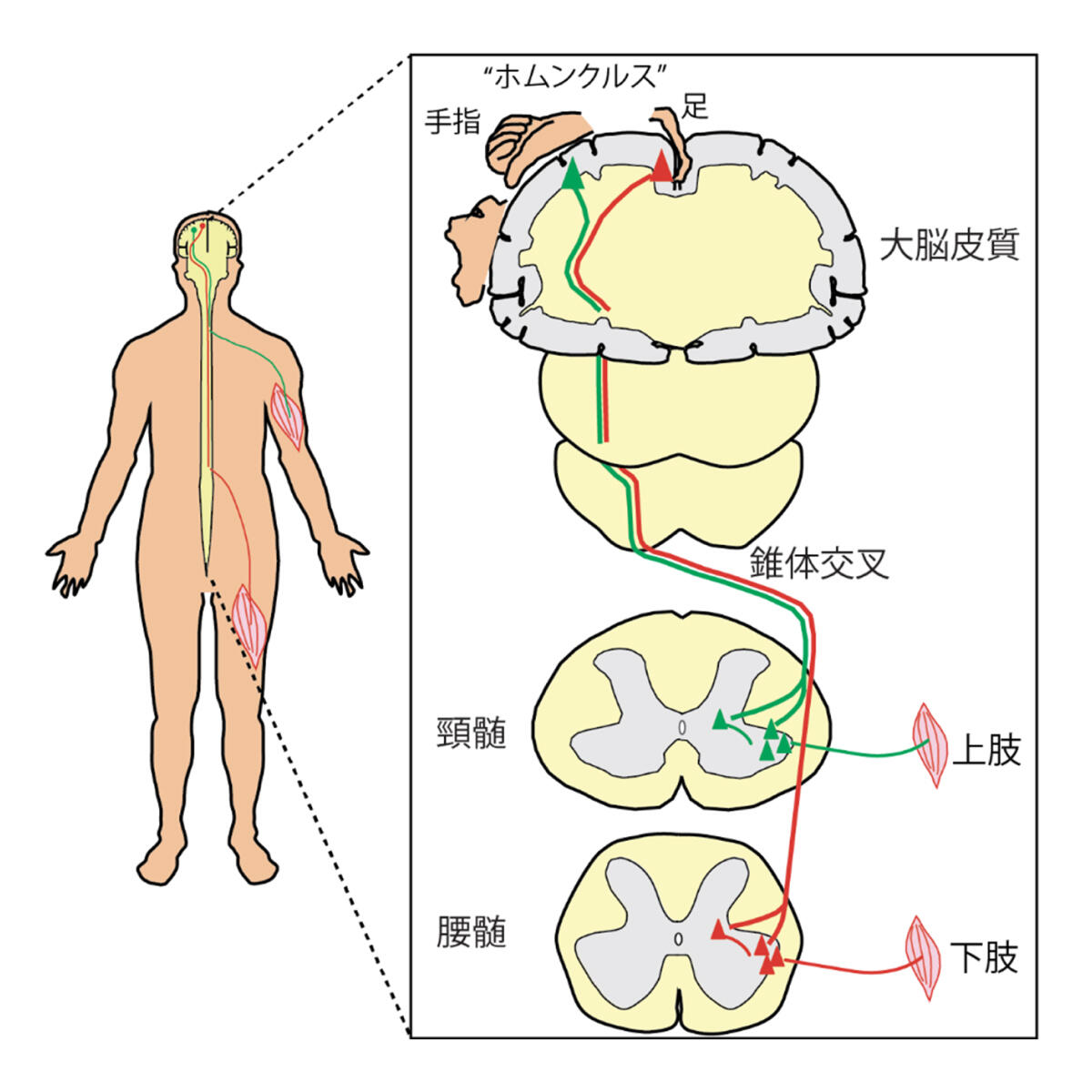
図2.運動を制御する皮質脊髄路
皮質脊髄路は,大脳皮質の運動を制御する領野(運動野など)の第V層にあるニューロンをもととする.このニューロンは,その軸索を,脳幹延髄の底部にある錐体へと伸ばし,そこでヒトではおおよそ80-90%が交叉し,反対側の脊髄へと伸ばす(図2).いわゆる,左の脳が右半身の運動を制御し,右が左を制御するもととなる回路である.錐体を通ることから,臨床では錐体路とよく呼ばれる.起始部である大脳皮質の運動野には,刺激すると各体部位を動かす領域があることが有名で,ラテン語で小人を意味するホムンクルスとも呼ばれる.そしてこの小人の各部位は,皮質脊髄路の投射様式が作り出していると考えられる.例えば,手指を動かす領域の皮質脊髄路は,頸髄の運動ニューロンへと伸び手指の筋へつながり,足を動かす領域は腰髄へ伸びて足の筋に至る,という具合だ.
さて,ホムンクルスはPenfieldらがヒトの脳で見出したことで有名であるが2,実際にはそこに至る長い歴史がある.皮質脊髄路の発見の歴史は,いくつかの本や総説に詳しい(図3)3, 4, 5, 6, 7.
思考の中枢が脳であるかどうかも定かでなかった5000年前のパピルスや紀元前のヒポクラテスの文書では,左頭部の障害が右半身を,右の障害が左半身を侵すことがすでに記され,交叉するシステムが示唆されているという.17-18世紀,脳に機能的な領野があることがまだわかっていない時代に,Boyle (1691) は,頭蓋骨折の骨片を除くと反対側の四肢の運動麻痺が治癒したことから,大脳に運動野の存在を推察した.解剖学的には,錐体交叉がイタリアのMistichelli(1709)やフランスの外科医,解剖学者Pourfour du Petit(1710)により記述されたが,この構造がどこからきているか同定されていない.皮質脊髄路の存在が浮き彫りになるのは19世紀に入ってからである.1810年,骨相学と大脳の機能局在説を広めたGallとSpurzheimは,錐体の起始部が大脳皮質から来ることを脳解剖で見ている.1851年,Türckはヒト脳の錐体の損傷で,脊髄に変性した神経経路が見られたことから,錐体が脊髄へつながること,また大脳や主に基底核からつながると推察した.Broca(1861)が大脳皮質言語中枢の損傷による失語症を報告し,機能局在説がさらに注目された後,Jackson(1864)は,大脳のてんかん発作で四肢が動く観察から,大脳皮質に運動の領野があると想定した.1870年には,ドイツのFritsch とHitzigの有名な実験がある.彼らはイヌの大脳皮質を電気刺激したところ,今日の運動野とよばれる場所の刺激で,反対側の四肢の運動を誘発することを発見した.ここから畳みかけるように発見は続く.翌年,Gudden(1872)はイヌの大脳皮質損傷後に変性した神経を観察し,錐体と脊髄が大脳皮質からつながることを報告,Ferrier(1873)は,霊長類を含めたいくつかの動物で運動を誘発する領域が大脳皮質の中心前回にあると同定した.Betz(1874)はこの領域に,巨大な細胞(Betz細胞)があることを見出し,運動の起始部と推定した.このあたりで,大脳皮質から錐体,脊髄へつながり運動を支配する経路の存在が,生理学,解剖学的に見えてきている.
一方当時,神経の経路は,神経細胞同士が融合して網状のネットワークを形成しているのか(網状説),神経細胞が個々に独立しシナプスを介して情報伝達しているのか(ニューロン説),論争のある時代であった.しかし,ゴルジ染色で神経回路の構造を克明に記載し,ニューロン説を確かにしたRamón y Cajalは,大脳皮質の中心前回にある細胞の形状を解析し,今日教科書で一般に見られる皮質脊髄路の構造を美しく描きだしている(1893)8.その後20世紀初頭には,CampbellやBrodmannがヒトの脳局在を解剖学的に分類し,中心前回にあたる4野が運動野,6野が運動前野などとした.こうした研究がつながり,Woolseyらはサルなど動物で運動野の体部位局在を見出し,ついに Penfield は,局所麻酔での脳外科手術中にヒト大脳を刺激し,ヒトホムンクルスの発見へつながって行くのである.
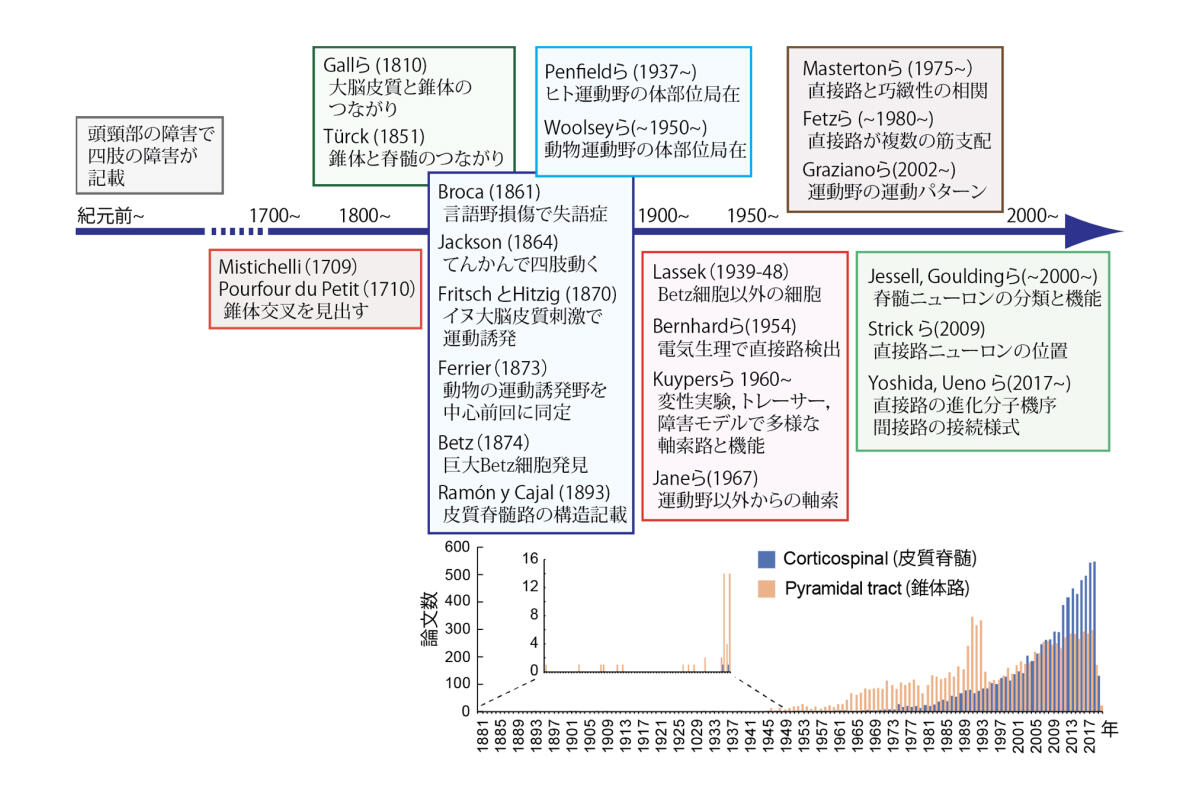
図3.皮質脊髄路の発見の歴史
右下:PubMedのキーワード検索による発表論文数の推移.PubMed上,錐体路(pyramidal tract)は1881年に現れ,皮質脊髄(corticospinal)の語は,Lassekの論文 (1946) からスタートしている. 錐体路は延髄に終わる線維も含むので,皮質脊髄路と呼ばれることが近年増えている.
さてこの皮質脊髄路の起始細胞は,運動野第V層にある巨大なBetz細胞と考えられていた.これはHolmesとMay(1909)により,皮質脊髄路を頸髄レベルで切断した場合に,逆行性に変性するのが巨大細胞のみであるという観察から支持された3.しかしLassek(1939-48)によって,皮質脊髄路はBetz細胞以外からも出ていると示唆された.ヒトの錐体において100万本の軸索が通っているのに対し,Betz細胞はおよそ34,000しかないという.したがって,錐体路中に占めるBetz細胞の線維数は3~4%にすぎないというのだ.実際,近年の組織学的解析においてもBetz細胞は第V層神経細胞の10%程度しかないという9.さらに,運動野を除いた患者において,40%の軸索が残っていたことから,運動野以外から出ている軸索があると考えられ(Jane et al., 1967),サルの実験でもそれが確かめられている (Russell and De Myer 1961).1970年代以降は,神経を変性させる古典的な実験から神経トレーサーの発展があり,各脳領域の神経細胞がどこへ軸索を伸ばしているのか,直接見て確かめられるようになる.特に,Kuypersらをはじめとした多くの研究者の報告から,皮質脊髄路が実は運動野からだけでなく,少なくとも9を超える大脳皮質の様々な領野から出ていることがわかってきている10, 11, 12.これらの観察結果は,皮質脊髄路が脊髄へ単に運動の指令を送るというものではなく,実際は,多様な情報を送っていると示唆している.
皮質脊髄路の中で,もっとも単純と考えられる経路は,大脳皮質運動野のニューロンの軸索が直接脊髄の運動ニューロンに接続するCorticomotoneuronal (CM) connection(直接路)と言われるものである4, 13, 14.この経路は,1950年代に電気生理学的に見いだされ15,80年代には解剖学的にも検出されている16.面白いことにこの接続は,高等な霊長類とヒトにしかない様式で,これは指の一本一本を独立して動かすなど手指の巧緻性を発揮する独自の経路と今日考えられている.直接路が,巧緻性の低いリスザルなどの霊長類では少なく,さらに他の哺乳類では顕著に見られなくなる,といった相関からも推測されている13.皮質脊髄路は,調べられている限り全ての哺乳類に存在し,進化の過程により大きく分けて4つの異なるタイプがあるという.それぞれの構造は巧緻性など各動物の行動様式に相関しているようである(図4)3, 4.
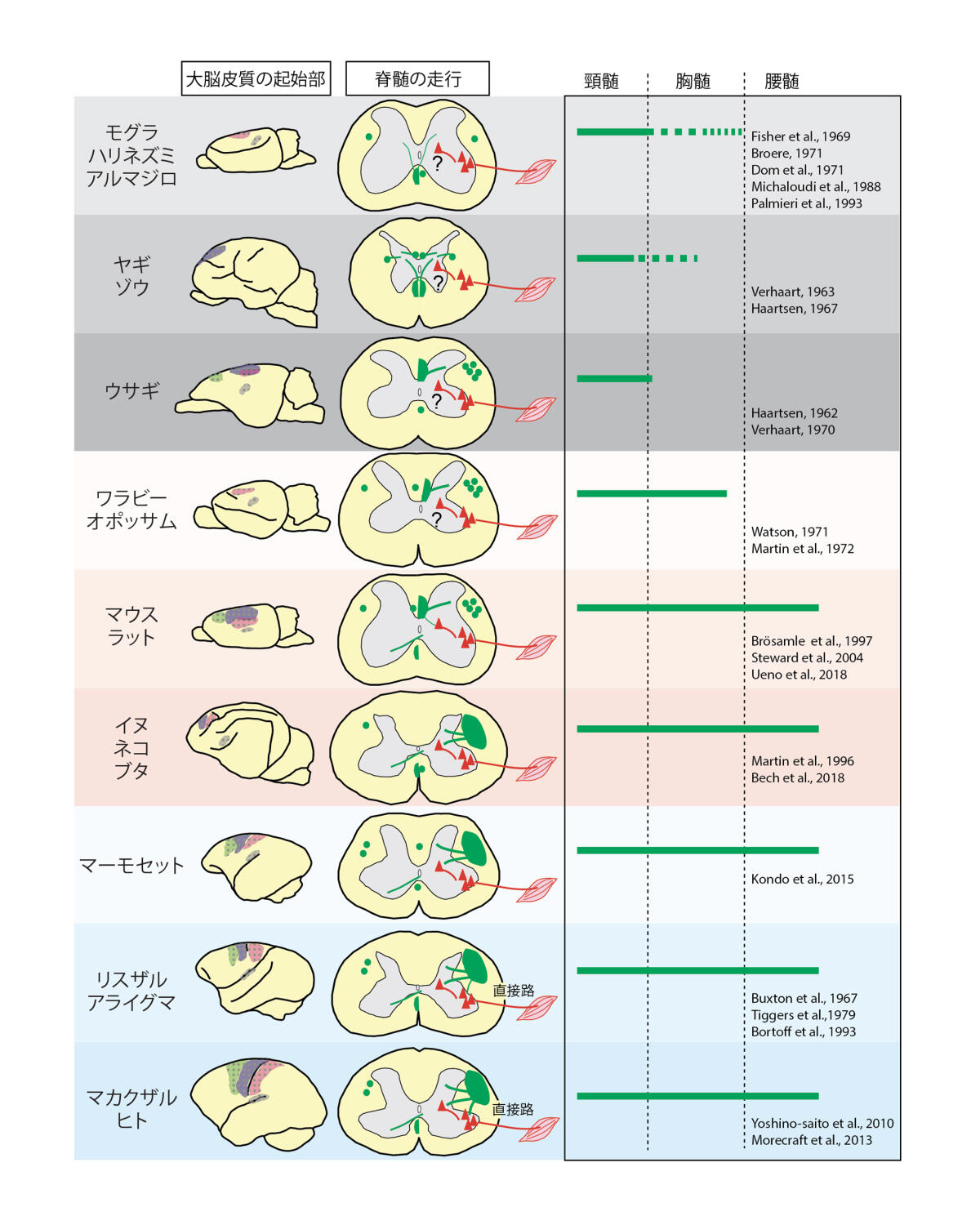
図4.動物種による皮質脊髄路の多様性
HeffnerとMasterton (1975, 1983)は,各動物の指の巧緻性を1から7に分類し,皮質脊髄路の構造との相関を見出した17, 18.進化的に古いものは,軸索が脊髄灰白質の主に背側へ投射する.背側は感覚情報が入力してくる位置であることから,皮質脊髄路はこの感覚情報をコントロールする回路であったとも推察されている3, 13, 19.一方,ヒトや高等な霊長類では,脊髄灰白質の腹側へ投射する線維が豊富で,一部遠位の筋肉の運動ニューロンに直接結合することから,個々の指など巧緻的な運動を可能にしていると想定される.筋肉への経シナプスウイルストレーサーによる実験から,この直接路ニューロンは運動野の一部領域に多く存在するが,一方他の領域には間接路ニューロンも多いことが示されている20.また直接路のニューロンはBetz細胞に限らないことも付記しておきたい21.したがって,ヒトにおいてもおそらく直接路ニューロン以外の多様性があるだろう.他方,モグラではそもそも皮質脊髄路の多くが交叉せず脊髄の腹側を通り,ウサギやヤギでは頸髄までしか皮質脊髄路が到達しない3, 10.われわれが実験でよく用いるげっ歯類では,ヒトで脊髄の背外側を通る軸索路が,背内側をメインに伸びている.種によってバラバラであるのは面白い.この投射様式の中に,各動物種固有の運動様式が垣間見え,動物の進化-神経の構造-行動様式の連関を映し出している興味深い回路である.例えば,発生の時期に軸索の伸長をガイドする因子の発現の違いによって,種間で直接路の有無と運動様式が決まると示されている22.
このように皮質脊髄路は,少なくとも19世紀後半にはその原型がわかり,その後,投射様式や大まかな機能がわかってきた.しかし,協調的な筋運動を生み出すからくり,すなわちどのような神経細胞の接続と活動によって,かくも複雑で絶妙な運動のパターンを生み出すことができるのか,動作原理はいまだ謎のままである.
大脳皮質と脊髄,それをつなぐ回路の中に,動作をコマンドする装置,すなわち神経回路の配線が備わっていることは確からしい.大脳皮質運動野には,運動の様々なパラメーター(運動の強度や方向)に反応する神経細胞群があり,各筋運動の強度と時空間的パターンを作り出していると考えられる(図5A)23, 24.実際,直接路のニューロンを刺激すると,ある特定の筋群の反応が起こり25,さらに運動野を長く刺激すると,手を口元へもってくるなど日常見られるまとまった運動のパターンが誘発される26, 27, 28.一方,脊髄では,離断実験などにより,反射や歩行を自動的に行う神経回路が備わっていることがよくわかっている.複数の筋肉の協調的な運動に連動する神経細胞の存在も示され,筋シナジーとも呼ばれる基本的な運動を生み出す配線が脊髄に存在すると考えられる29, 30.以上から,大脳皮質や脊髄の中に,運動パターンを生み出せる回路が内在しているとわかる.
では,こうしたコマンドシステムを持つ大脳皮質と脊髄の回路は,お互いどうつながり連関しているのであろうか.大脳皮質からの指令は,様々なバイパスを通り脊髄へ到達することが知られる(図1).Kuypers らは,脳各部の傷害実験から,それぞれの役割が違うことを示唆した.皮質脊髄路はより四肢末端の巧緻的な運動に大事で,一方,網様体など脳幹部を通って脊髄へ至る経路は,姿勢保持などより体幹に近いものを制御すると考察されている31, 32.ヒトが錐体路を障害された場合には,ごく初期の弛緩性の麻痺から痙性の麻痺に移っていく.このことから皮質脊髄路は,運動の指令だけでなく,各種筋群の緊張や協調的な動きを制御しているとうかがえる.わかっていないのは,運動パターンを備える大脳皮質や脊髄がどのようなつながりと仕組みで連動しているのかということである(図5A).大脳皮質の様々な領野から出る皮質脊髄路もこのつながりに関わっているであろうが,その仕組みもよくわからない.皮質脊髄路に残された大きな謎といえる.
わたしたちはげっ歯類であるマウスを用いて,皮質脊髄路に残された謎,特にそのつながり方や機能の解明に取り組んできた(図5B)33.マウスでは,遺伝子改変技術とそれに付随する神経の標識や操作法が強みを発揮する.近年特に,特定の神経細胞のつながりや活動を観察したり,その活動を上げ下げ操作することで,神経細胞種レベルでの機能がわかるようになってきた.皮質脊髄路に例を見るように,古くから知られていても,技術的に解決できなかったような問題が,新たに掘り起こされ解決できる時代になっている.脊髄でいえば,90年代以降JessellやGouldingを中心に,脊髄の多様なニューロン種が同定され34,その細胞を操作することで,運動や各種感覚の伝達や抑制に関わる多様な働きが分かってきた.こうした流れがあるのだが,一方,脊髄の多様なニューロンが上位中枢とどうつながり機能しているのかよくわかっていない.
わたしたちはまず,マウスの皮質脊髄路においても,運動野だけでなく,感覚情報を受け取る感覚野など複数の大脳皮質領域から,異なる投射様式の軸索が皮質脊髄路を通って脊髄へ伸びていることを見出した(図5B)33.面白いのは,運動野のニューロンは,最終的に筋肉へとつながる一方,感覚野から筋肉へのつながりは悪いということで,運動に直接関与する経路と,間接的に寄与する経路があることを示唆する.実際これらの回路は,それぞれ脊髄内で運動を伝達するのみならず,感覚情報を伝達したり遮断するなど,異なる機能に関わるニューロンと接続することがわかった.そこで,それぞれの回路や神経細胞を実験的に遮断してみると,運動の最中に,腕が伸ばせなかったり,到達前に腕を引っ込めたり,違う方向に腕を伸ばしたり,ものを掴めなかったり,掴んだものを離せなかったり,と多様な障害の種類が現れた.したがって,皮質脊髄路には,運動の出力や方向などの性質,運動中に感ずる感覚の制御など,動作の最中の異なる要素を制御する回路が内在していると考えられた.このような多様性は,皮質脊髄路のニューロンが,運動中の異なるフェーズで実際に活動するという報告からも示唆される35.運動では,つかんだ取手や鍵盤の硬さなど微妙な感覚の情報や,腕の位置や筋の収縮など自らの体位置の情報をうまくとらえ,運動の位置や強度の調整を行う必要がある.よって運動情報の指令だけでなく,こうした感覚情報の制御も皮質脊髄路が行っていると考えられる36.神経細胞種レベルでの皮質脊髄路の多様な接続と機能は,特定の神経細胞の標識や操作ができるマウスで見えてきたことである.
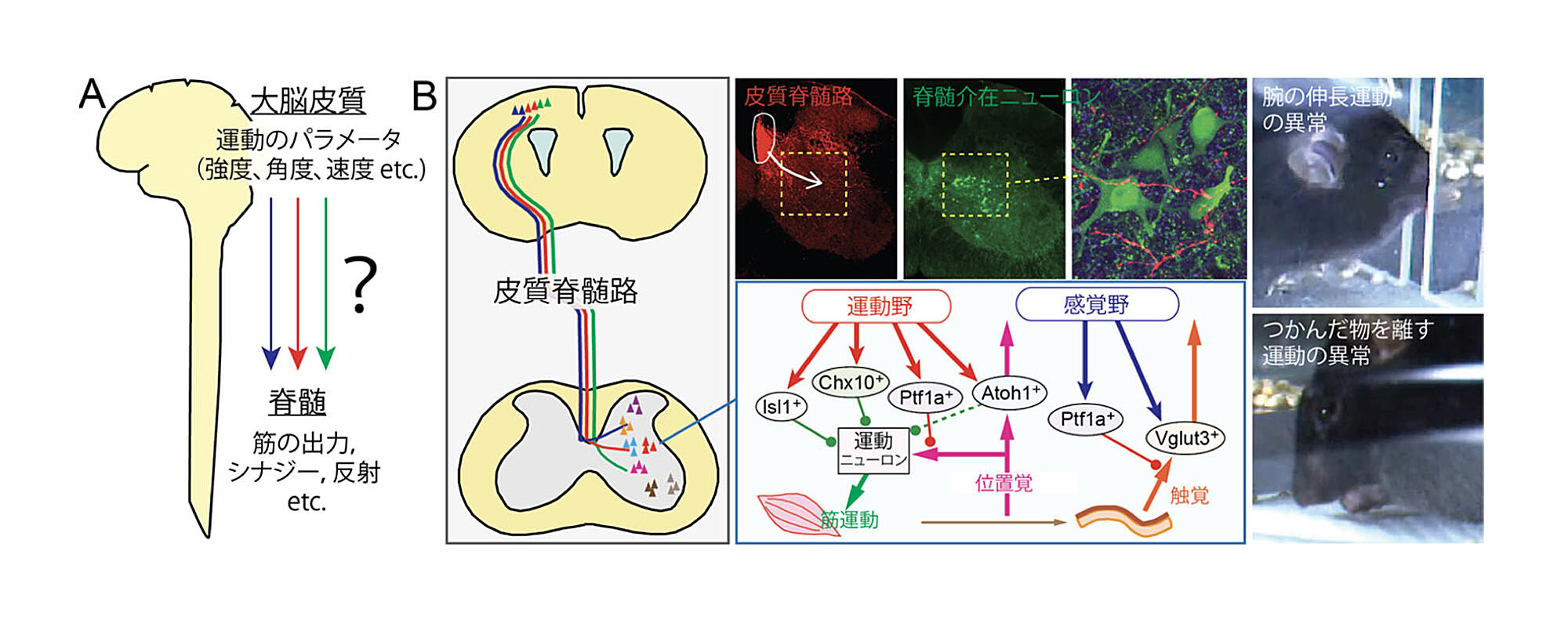
図5.マウスの多様な皮質脊髄路33
(B)マウスでは運動野,感覚野などから脊髄へ異なる投射があり,筋へつながるもの,つながらないものがある(左).
これらは脊髄で,運動,感覚に関わる多様なニューロンと接続する(中).
各回路の遮断で,運動中に多様な障害が現れる(右).
皮質脊髄路が,運動の指令,出力,感覚の制御など,多様な接続と機能を持つことを示唆する.
動きを生み出す脳の仕組みの理解は,これからさらに進むであろう.特に機能要素ごとの回路は興味深い.例えばマウスでは,感覚野から伸びる顕著な経路や,脊髄運動介在ニューロンにつながる経路が見られた.近年ヒトにおいてもトラクトグラフィーにより多様な大脳皮質領域から伸びる皮質脊髄路の経路が見出されている37.また直接路だけでなく,脊髄介在ニューロンを介した動作が,巧緻性の高いサルで重要であることが示されている14.種間の違いがよく指摘されるのだが,原型として保存された回路もあるのではなかろうか.一方,よくわかっていないことも多い.例えば,屈筋と伸筋など数ある筋肉を,数ある動作パターンで協調的に制御できる回路の仕組みは不明だ.今後,各運動脳領域あるいはその間の回路の構造を詳細にし,各神経細胞の活動のパターンをオンラインで解読できれば,動作モデルが構築されていくだろうが,動きを生み出す真の原理の解明はまだ先となるかもしれない.
最後に,脳卒中や脳脊髄の損傷,ALSなど,皮質脊髄路の脱落で運動機能が障害される疾患においては,健常時の回路の姿を見出すことで,どのような回路の再建が必要であるかが見えてくる.今時点でも,そう単純でない回路の再建方法が求められることが容易に理解できる.今後,神経回路の再生技術やリハビリテーション,あるいはBMIによる人工回線などを用いて,さらに見いだされていくであろう神経回路の本当の姿をいかにして再建できるかということが,次の100年の大きな課題として続いていくと考えられる.
文献
- Grillner S. Fundamental of motor systems. In: Fundamental Neuroscience (eds Squire LR et al.). Academic Press (2003).
- Penfield W, Boldrey E. Somatic motor and sensory representation in the cerebral cortex of man studied by electrical stimulation. Brain 60, 389-443 (1937).
- Armand J. The origin, course and terminations of corticospinal fibers in various mammals. Prog Brain Res 57, 329-60 (1982).
- Porter R, Lemon R. Corticospinal function and voluntary movement. Oxford university press (1993).
- Kandel ER, Schwartz JH, Jessell TM, Siegelbaum SA, Hudspeth AJ. Principles of Neural Science. McGraw-Hill Companies (2013).
- Swanson. LW. Neuroanatomical terminology: a lexicon of classical origins and historical foundations. Oxford university press (2015).
- 萬年甫. 頭のなかをのぞく神経解剖学入門. 中山書店 (2013).
- Swanson LW, Lichtman JW. From Cajal to connectome and beyond. Annu Rev Neurosci 39, 197-216 (2016).
- Rivara CB, Sherwood CC, Bouras C, Hof PR. Stereologic characterization and spatial distribution patterns of Betz cells in the human primary motor cortex. Anat Rec A Discov Mol Cell Evol Biol 270, 137-51 (2003).
- Kuypers HG. Anatomy of the descending pathways. In: The Nervous System, Handbook of Physiology (eds Brooks V). Williams and Wilkins (1981).
- Darian-Smith I, Galea MP, Darian-Smith C, Sugitani M, Tan A, Burman K. The anatomy of manual dexterity. The new connectivity of the primate sensorimotor thalamus and cerebral cortex. Adv Anat Embryol Cell Biol 133, 1-140 (1996).
- Darian-Smith C, Lilak A, Garner J, Irvine KA. Corticospinal sprouting differs according to spinal injury location and cortical origin in macaque monkeys. J Neurosci 34, 12267-79 (2014).
- Lemon RN. Descending pathways in motor control. Annu Rev Neurosci 31, 195-218 (2008).
- Isa T. Dexterous hand movements and their recovery after central nervous system injury. Annu Rev Neurosci 42, 315-35 (2019).
- Bernhard CG, Bohm E. Monosynaptic corticospinal activation of fore limb motoneurones in monkeys (Macaca mulatta). Acta Physiologica Scandinavica 31, 104-12 (1954).
- Shinoda Y, Yokota J, Futami T. Divergent projection of individual corticospinal axons to motoneurons of multiple muscles in the monkey. Neurosci Lett 23, 7-12 (1981).
- Iwaniuk AN, Pellis SM, Whishaw IQ. Is digital dexterity really related to corticospinal projections?: a re-analysis of the Heffner and Masterton data set using modern comparative statistics. Behav Brain Res 101, 173-87 (1999).
- Yoshida Y, Isa T. Neural and genetic basis of dexterous hand movements. Curr Opin Neurobiol 52, 25-32 (2018).
- Lemon RN, Griffiths J. Comparing the function of the corticospinal system in different species: organizational differences for motor specialization? Muscle Nerve 32, 261-79 (2005).
- Rathelot JA, Strick PL. Subdivisions of primary motor cortex based on cortico-motoneuronal cells. Proc Natl Acad Sci U S A 106, 918-923 (2009).
- Lemon R. Recent advances in our understanding of the primate corticospinal system. F1000Research 8:274 (2019).
- Gu Z, et al. Control of species-dependent cortico-motoneuronal connections underlying manual dexterity. Science 357, 400-4 (2017).
- Kalaska JF, Rizzolatti G. Voluntary movement: the primary motor cortex. In: Principles of Neural Science (eds Kandel ER et al.). McGraw-Hill Companies (2013).
- Levine AJ, Lewallen KA, Pfaff SL. Spatial organization of cortical and spinal neurons controlling motor behavior. Curr Opin Neurobiol 22, 812-21 (2012).
- Fetz EE, Cheney PD. Postspike facilitation of forelimb muscle activity by primate corticomotoneuronal cells. J Neurophysiol 44, 751-72 (1980).
- Graziano MS, Aflalo TN. Mapping behavioral repertoire onto the cortex. Neuron 56, 239-51 (2007).
- Ramanathan D, Conner JM, Tuszynski MH. A form of motor cortical plasticity that correlates with recovery of function after brain injury. Proc Natl Acad Sci U S A 103, 11370-5 (2006).
- Harrison TC, Ayling OG, Murphy TH. Distinct cortical circuit mechanisms for complex forelimb movement and motor map topography. Neuron 74, 397-409 (2012).
- Takei T, Confais J, Tomatsu S, Oya T, Seki K. Neural basis for hand muscle synergies in the primate spinal cord. Proc Natl Acad Sci U S A 114, 8643-8 (2017).
- Levine AJ, et al. Identification of a cellular node for motor control pathways. Nat Neurosci 17, 586-93 (2014).
- Lawrence DG, Kuypers HG. The functional organization of the motor system in the monkey. I. The effects of bilateral pyramidal lesions. Brain 91, 1-14 (1968).
- Lawrence DG, Kuypers HG. The functional organization of the motor system in the monkey. II. The effects of lesions of the descending brain-stem pathways. Brain 91, 15-36 (1968).
- Ueno M, et al. Corticospinal circuits from the sensory and motor cortices differentially regulate skilled movements through distinct spinal interneurons. Cell Rep 23, 1286-300. (2018).
- Alaynick WA, Jessell TM, Pfaff SL. SnapShot: spinal cord development. Cell 146, 178 (2011).
- Wang X, et al. Deconstruction of corticospinal circuits for goal-directed motor skills. Cell 171, 440-55.e414 (2017).
- Liu Y, et al. Touch and tactile neuropathic pain sensitivity are set by corticospinal projections. Nature 561, 547-50 (2018).
- Chenot Q, et al. A population-based atlas of the human pyramidal tract in 410 healthy participants. Brain Struct Funct 224, 599-612 (2018).
- Nudo RJ, Masterton RB. Descending pathways to the spinal cord, III: Sites of origin of the corticospinal tract. J Comp Neurol 296, 559-83 (1990).
