



(2016年6月7日執筆)
担当:塚野浩明先生
所属:基礎神経科学部門
システム脳生理学分野
1.大脳聴覚野
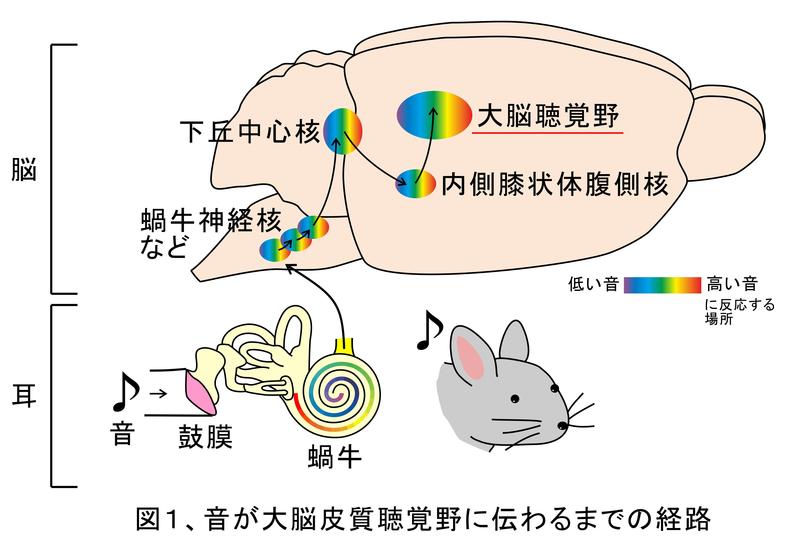
私たちは音を通じてさまざまな外界の状況を認知しています。例えば、電話の相手が誰なのかが声だけで判ったり、後ろから車が近付いてきたことを知ることが出来ます。
音はまず耳に入りますが、実際に音を認識しているのは脳です。音は耳に入ると蝸牛(かぎゅう)に伝わります(図1)。蝸牛の渦巻いた管の中には音によって振動する基底板という板が伸びており、音が低くなるにつれて振動する場所が少しずつ奥にずれていく特徴があります。これにより、音の高さ情報は1枚の板上の場所情報に変換されます。この音の高低の分布はトノトピーと呼ばれ、神経を伝わって大脳の「聴覚野」と呼ばれる領域に辿りつくまで保存されます。高さは重要な音の要素の1つではありますが、この様に高さは耳の時点でかなり精密に分析されています。したがって、音を伝える系路の最初の方で高さの情報処理は終わっていることが想像されます。実際、動物が単純な音の高さを知覚するには聴覚野は必要ないことが過去の研究で判っています1)。つまり、音情報が最後に到達する聴覚野はもっと複雑で高度な音成分を処理しているはずなのです。
2.機能局在
しかし、大脳聴覚野がどのような音をどのような神経メカニズムで情報処理しているのか、ほとんど判っていません。研究が進んでいないのは、聴覚野で処理している音成分自体が複雑で研究が難しいこともあります。しかしそれ以前の問題として、マウス聴覚野の構造自体が最近までよくわかっていなかったということが大きいと考えます。
脳の表面は、外から見ると肌色で一見のっぺりとしています。しかし、脳は見えない多くの領域に分かれていて、それぞれ異なった機能を持っていると考えられています。例えば、音を処理する聴覚野、目で見た情報を処理する視覚野、食べたものの味を処理する味覚野、などの領域に分かれています。この様に脳の場所によって担う機能が異なる原理を「大脳皮質の機能局在」といい、現在の脳科学の前提となっています。これらの脳領域はさらに多くの小さな領域に分かれています。大脳聴覚野も単一の脳部位でなく、いくつかの小領域の集合体なのです。機能局在論に鑑みると、小領域もそれぞれ異なった機能を担っているはずです。したがって、聴覚野の機能を解明するためには、聴覚野を構成するそれぞれの小領域の機能を知る必要があります。そして、各小領域から正確に神経活動を記録し研究するために、各小領域の場所を正しく把握しなければならないのは当然です。聴覚野の小領域の場所を描いたものを聴覚野地図(聴覚野マップ)と呼びます。マウスを用いると幅広い解析を行うことが出来るので、現在の脳科学では特にマウスの聴覚野マップを正確に描出することが必要不可欠なのです。
3.新しい聴覚野マップ
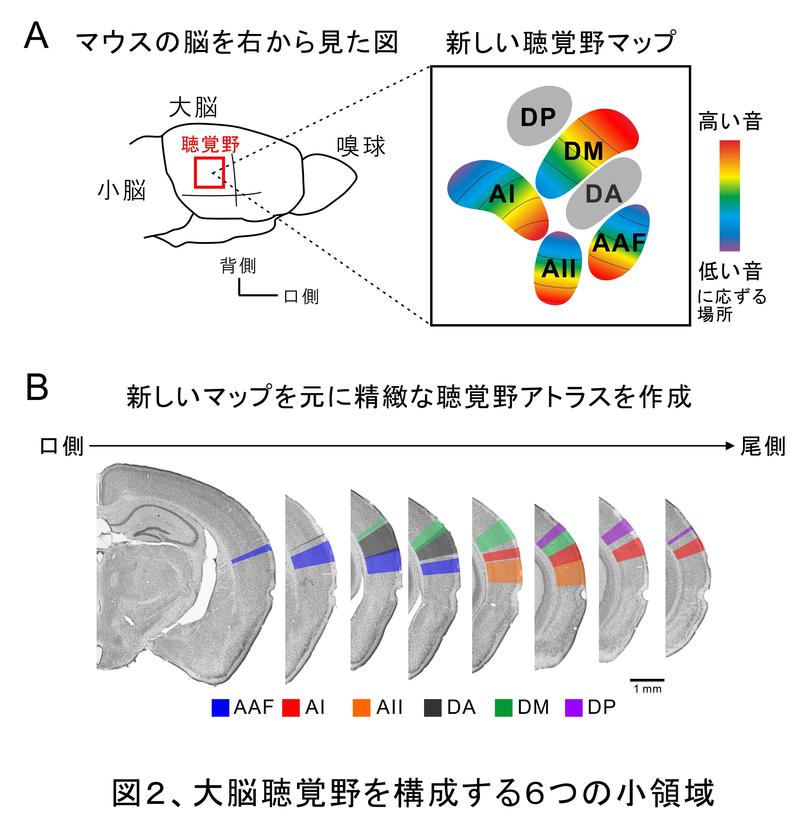
マウスはとても小さい動物で、脳全体がヒトの小指の先くらいの大きさしかありません。聴覚野になると全体で2 × 2 mmくらいしかないため、精密に観察することが難しいのです。したがって、これまで出版・公開されているマウス聴覚野マップはとても大雑把なものでした。しかし、ある領域Aの機能を炙り出したいのに、領域Aと別の領域Bとごちゃまぜにして研究していては、いつまでたってもきちんとしたデータを出すことは不可能です。
我々は、小さい範囲を観察することに長けている光を用いることで脳活動を捉えるフラビン蛋白蛍光イメージングという方法でマウス聴覚野が音に反応する様子を観察しました(詳しくは、脳研コラム『マウス大脳皮質感覚野の経頭蓋フラビン蛋白蛍光イメージング』を参照)。すると、6個の音に反応する場所が見つかりました(図2)2,3)。私たちはさらに、音に反応した6つの場所の細胞の構造を調べました。すると6つの場所はそれぞれ異なる特徴を持つ細胞から作られていることが判りました4)。これによりマウス聴覚野は少なくとも6つの小領域から成り立っていることが判りました。現在、7つ目8つ目の小領域も発見しています(未出版)。
4.新しい聴覚伝導路の姿
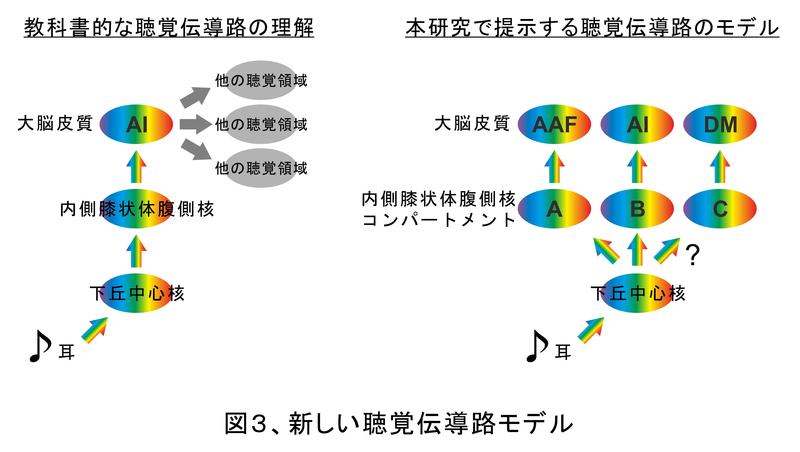
聴覚野は音情報が最後に到達する所と書きました。聴覚野の一つ手前の脳部位は内側膝状体腹側核(ないそくしつじょうたいふくそくかく)という名前が付いています(図1・図3)。少し複雑な名前ですが、ヒトの脳では膝のように曲がった恰好をしている場所の内側にあって、さらに少し腹側にある神経核なので、このような名前がついています。聴覚野もよく判っていませんが、実は内側膝状体腹側核もその全貌がよく判っていません。というのは、聴覚野は脳の表面にありますが、内側膝状体腹側核は脳の内側の方に位置しているため観察がさらに難しいからです。私たちは聴覚野のそれぞれの小領域が内側膝状体腹側核からどのような系路で情報を受け取っているか調べようと思いました。そこで、神経トレーサーをいう試薬を各小領域に少し注入しました。神経トレーサーは神経細胞の中を運ばれて行く性質があります。つまり、それぞれの小領域が内側膝状体腹側核のどこと繋がっているのか細かく判ります。その実験の結果、聴覚野の各小領域は内側膝状体腹側核の中でそれぞれ異なる場所と繋がっていることが判りました(図3)6,7)。同様の知見が国内の他の研究室からもほぼ同時期に報告されています8)。この事実は、内側膝状体腹側核がいくつかのコンパートメントに分かれていて、それぞれのコンパートメントから1対1に音情報を受け取っていることを意味しています。この様なきれいなコンパートメント構造があることは、最近まで見つかっていませんでした。
これらの事実は重要なことを示唆しています。従来は、内側膝状体腹側核から大脳聴覚野に音情報がまず入る玄関は一次聴覚野(AI)という小領域で、そこから他の小領域に情報別に送られると考えられていました(図3)。しかし、内側膝状体腹側核の時点で複数のコンパートメントに分かれていて、そこから聴覚野の各小領域に行く系路も別々ということは、各系路が別々の音成分を運んでいる可能性が高くなります。そうなると、聴覚野のどの小領域にどの音成分を送るのかを選択して送り分ける機能が内側膝状体腹側核の時点で存在すると推測されます。内側膝状体腹側核は単にバトンを渡すだけでなく積極的に聴覚情報処理に参加している可能性が高くなりました。過去に別の動物種で行われた研究でもこれを推測させるような研究がありましたが10,11,12)、それがさらに確からしくなりました。
内側膝状体腹側核の一つ手前の脳部位は、下丘中心核(かきゅうちゅうしんかく)という所です。現在は下丘中心核は単一の構造だと考えられています(図3)。しかし、内側膝状体腹側核がいくつかのコンパートメントに分かれているならば、下丘中心核もいくつかのコンパートメントに分かれているのかもしれません。もしくは、一つの下丘中心核から複数の内側膝状体腹側核コンパートメントに分岐しているのかもしれません。聴覚伝導路の始まりに近い方がどのようになっているのかを知ることは聴覚情報の振り分けがどのようになされているか知るために重要ですので、いずれ研究されることと思います。いずれにしても、従来考えられていたより複雑な聴覚伝導路によって聴覚機能が実現しているということが判りました。
5.大脳聴覚野研究の今後の展望
前述したように、聴覚野は単純な音の高さを知覚することには必要ありません。では、聴覚野を必要とするようなもっと複雑で動物にとって大切な音とは何でしょうか。それは鳴き声だと思います。鳴き声によって他の個体とコミュニケーションが取れることで、雄雌間コミュニケーションや母仔間コミュニケーションが実現し、動物が生存したり子孫が残せたりします。またヒトではこれらの神経基盤を転用して音楽を楽しむことにも使われている可能性も高いと考えます。これらの音処理に、聴覚野小領域がどのように相互作用しているのか、精細に可視化して行きたいと思っています。
参考文献
1. Ono, K., Kudoh, M. & Shibuki, K. Roles of the auditory cortex in discrimination learning by rats. Eur. J. Neurosci. 23, 1623-1632 (2006).
2. Tsukano, H., Horie, M., Hishida, R. & Shibuki, K. New subarea in the rostrodorsal part of the primary auditory cortex in mice. J. Physiol. Sci. 63, S205 (2013).
3. Tsukano, H., Horie, M., Bo, T., Uchimura, A., Hishida, R., Kudoh, M., Takahashi, K., Takebayashi, H. & Shibuki, K. Delineation of a frequency-organized region isolated from the mouse primary auditory cortex. J. Neurophysiol. 113, 2900-2920 (2015).
4. Tsukano, H., Horie, M., Hishida, R., Takahashi, K., Takebayashi, H. & Shibuki K. Quantitative map of multiple auditory cortical regions with a stereotaxic fine-scale atlas of the mouse brain. Sci. Rep. 6, 22315 (2016).
5. Horie, M., Tsukano, H., Takebayashi, H. & Shibuki, K. Specific distribution of non-phosphorylated neurofilaments characterizing each subfield in the mouse auditory cortex. Neurosci. Lett. 606, 182-187 (2015).
6. Honma, Y., Tsukano, H., Horie, M., Ohshima, S., Tohmi, M., Kubota, Y., Takahashi, K., Hishida, R., Takahashi, S. & Shibuki, K. Auditory cortical areas activated by slow frequency-modulated sounds in mice. PLoS ONE 8, e68113 (2013).
7. Horie, M., Tsukano, H., Hishida, R., Takebayashi, H. & Shibuki, K. Dual compartments of the ventral division of the medial geniculate body projecting to the core region of the auditory cortex in C57BL/6 mice. Neurosci. Res. 76, 207-212 (2013).
8. Takemoto, M., Hasegawa, K., Nishimura, M. & Song, W.J. The insular auditory field receives input from the lemniscal subdivision of the auditory thalamus in mice. J. Comp. Neurol. 522, 1373-1389 (2014).
9. Storace, D.A., Higgins, N.C., Chikar, J.A., Oliver, D.L. & Read, H.L. Gene expression identifies distinct ascending glutamatergic pathways to frequency-organized auditory cortex in the rat brain. J. Neurosci. 32, 15759-15768 (2012).
10. Storace, D.A., Higgins, N.C. & Read, H.L. Thalamic label patterns suggest primary and ventral auditory fields are distinct core regions. J. Comp. Neurol. 518, 1630-1646 (2010).
11. Lee, C.C., Imaizumi, K., Schreiner, C.E. & Winer, J.A. Concurrent tonotopic processing streams in auditory cortex. Cereb. Cortex 14, 441-451 (2004).
12. Lee, C.C., Schreiner, C.E., Imaizumi, K. & Winer, J.A. Tonotopic and heterotopic projection systems in physiologically defined auditory cortex. Neuroscience 128, 871-887 (2004).
研究分野
- システム脳生理学分野
