



I.概略
我々はマウス大脳皮質感覚野の神経活動を経頭蓋フラビン蛋白蛍光イメージングという方法で解析している。脳活動は酸素代謝の亢進を起こし、ミトコンドリア電子伝達系のフラビン蛋白を酸化型にする。酸化型のフラビン蛋白は緑色自家蛍光を発するので、この内因性蛍光シグナルを用いた脳活動のイメージングが可能となる(図1)。マウスでは頭皮を切開するだけで透明な頭蓋骨越しに大脳皮質を観察できるため、内因性フラビン蛋白蛍光シグナルを用いた大脳皮質感覚野活動の経頭蓋イメージングが可能である。マウス大脳皮質感覚野の脳活動を解析すると、どのような事が判るのだろうか?
大脳皮質感覚野は幼若期の経験により様々な経験依存的な修飾を受け、完成する。経験依存的修飾の分子・細胞メカニズムは盛んに解析が進んでいる。特に様々な遺伝子の働きを抑えたり、亢進したりする改変操作を行った動物系統は、分子機構の解明に大きな威力を発揮する。しかし遺伝子改変操作が既に確立しているマウスの高次脳機能の解析は進んでおらず、一方高次脳機能が詳しく解析されている霊長類の遺伝子改変操作は困難である。この状況を乗り越える方法には、マウスの脳機能を詳細に解析する方法を開発するか、霊長類などにおける遺伝子操作を容易に行う方法を開発するか、二つの対応が考えられる。我々は前者の方針に基づいて、経頭蓋フラビン蛋白蛍光イメージングによってマウスの皮質活動を可視化し、マウス視覚野、聴覚野、体性感覚野の経験依存的可塑性を含む高次脳機能の解析に取り組んでいる。
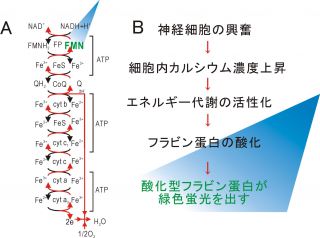
図1 フラビン蛋白蛍光イメージングの原理
A)ミトコンドリア電子伝達系とフラビン蛋白。フラビン蛋白は還元状態(FMNH2)と酸化状態 (FMN)を往来することで電子伝達を行う。酸化状態のフラビンのみが青い励起光の下で緑色の蛍光を発する。B)神経活動と内因性の緑色蛍光。神経が活動 すると細胞内のカルシウム濃度が高まり、ミトコンドリアの酸素代謝が高まってフラビン蛋白が酸化型にシフトする。その結果内因性の緑色蛍光が強まるので、 これを利用して脳活動のイメージングが出来る。
II.感覚野の古典的経験依存的可塑性
大脳皮質感覚野は視床を介する感覚情報を処理する脳部位である。五感それぞれに特化した感覚野が存在するが、経頭蓋的に容易に解析できるのは、視覚野、聴覚野、体性感覚野の三種類である。我々は経頭蓋フラビン蛋白蛍光イメージングでこの三種類の感覚野における経験依存的可塑性を解析し、今まで知られている古典的な現象が記録できることを確認した。
まず視覚野で古くから経験依存的可塑性として知られているものに、単眼遮蔽による眼優位性可塑性がある。これは生後の臨界期(マウスの場合4週齢前後)に左右の眼からの入力にアンバランスがあると、劣位眼に対する視覚野応答が抑圧される現象である。この眼優位性可塑性は、まずネコやサルなどの両眼視をする動物で研究されてきたが、実はマウス視覚野にもかなり大きな両眼視領域(両方の眼からの入力を重複して受ける脳部位)があり、良い実験対象であることが示されている。それ以来、マウスのメリットが生かされて、詳しい分子機構の解明が進んでいる。我々は経頭蓋フラビン蛋白蛍光イメージングを用いて単眼遮蔽による眼優位性可塑性の解析を行うことに成功した。又この実験系を用いて、生後1ヶ月くらいの時点に臨界期が存在することも確認できた。経頭蓋イメージングの大きなメリットの一つは、頭蓋骨の表面を透明な樹脂で保護すると感染や物理的ダメージを防止できるので、同一の動物から一定の期間を隔てて記録を繰り返すことができる点である。この特徴を利用して、単眼遮蔽の前後で同一のマウスから視覚野応答を記録したところ、遮蔽眼の応答が抑圧されるのに対し、健常眼の応答が増強されることが判った(図2A)。
一方、聴覚野の経験依存的可塑性のうち、最も単純なものは、特定の高さの音に曝して動物を飼育すると、その高さの音を特徴周波数(最も低い音圧で応答を起こす音の周波数)とするニューロンの数が増える現象である。この現象が経頭蓋フラビン蛋白蛍光イメージングにより可視化出来るかどうかを我々は試みた。その結果、特定の高さの音に曝して動物を飼育すると、その高さの音に対する蛍光応答が増強することを見いだした(図2B)。また、電気生理学的解析で明らかになっている臨界期が見られるかどうかもチェックしたが、少なくとも生後6週以前には明確な臨界期が存在しないことが判った。様々な環境音下で飼育した動物から聴覚野スライス標本を作製し、皮質の比較的浅い層を電気刺激してその反応をフラビン蛋白蛍光イメージングによって調べたところ、なるべく音を遮断して育てた動物では、皮質縦方向の興奮の広がりが小さいことが判った。即ち皮質縦方向のシナプス結合が音に曝して飼育することによって強化され、この可塑性が丸ごとの動物における聴覚野の経験依存的可塑性の基礎メカニズムとなっていることが推測される。
皮膚からの感覚情報を処理する体性感覚野の可塑性としてよく知られているものに、末梢入力の一部が遮断されると、対応する脳部位が周辺の末梢入力によって興奮するという現象がある。マウスの尾の一部を切断すると、これに応ずるはずの体性感覚野の一部が、尾の根元の部分の刺激によって応ずるようになることも判った。
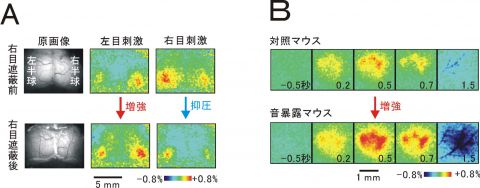
図2 経験依存的可塑性のイメージング
A)単眼遮蔽による視覚野可塑性。まず無処置の若いマウスの視覚野活動をイメージングで記録する(上段)。その後4日間右目の視野を遮断し、同じマウスで再び視覚野活動を記録する。視野が遮断されていなかった左目を介する視覚野応答が増強されるのに対し、視野が遮断されていた右目を介する応答は抑圧される(下段)。B)音暴露による聴覚野応答の増強。無処置マウスに10キロヘルツの音を聴かせると、聴覚野の応答が生ずる(上段)。各パネルの数字は音刺激の開始時点からの秒数。10キロヘルツの音を聴かせ続けながら飼育したマウスでは、10キロヘルツの音刺激に対する聴覚野の応答が増強している(下段)。一方、5キロヘルツや20キロヘルツの音に対する聴覚野応答はそれ程変化しない。
III.経験依存的可塑性と多彩な脳機能
経験依存的可塑性は古典的に知られている現象だけでなく、実は様々な高次脳機能の基礎メカニズムになっている。経験依存的可塑性を追求していくことで、これと密接に関連した高次機能を解明することができる。
経頭蓋蛍光イメージングによって、一次感覚野の脳活動だけでなく、高次感覚連合野の機能も明らかになってきた。高次感覚連合野の機能の一つは異なるモダリティの感覚情報を統合することである。例えば霊長類の頭頂連合野では体性感覚情報と視覚情報が統合され、空間認知がなされることはよく知られている。マウスではヒゲ情報と視覚情報の双方に基づいて空間認知がなされるので、霊長類の頭頂連合野に相当する部位でこの両者が統合されている可能性がある(図3)。マウスの眼前にプリズム眼鏡を装着すると、プリズムによって視覚情報に狂いが生ずるので、ヒゲ情報と視覚情報の食い違いが生する。このとき視覚野の応答が抑圧されるという経験依存的可塑性が生ずることが判った。この抑圧は、一見単眼遮蔽による抑圧と類似しているが、両眼からの入力が重複しない単眼視領域でも生じる。即ち単眼遮蔽による抑圧とは異なり、両眼からの入力が競合することによって抑圧が生ずるのではない。さらに予めヒゲを切ったマウスにプリズム眼鏡を装着させても抑圧は生じなかった。即ちヒゲを切っておくことによってヒゲからの情報が入らなければヒゲ情報と視覚情報の食い違いが検出されず、抑圧も生じない。一方単眼遮蔽による抑圧はヒゲを切っておいても通常と変わりなく生じた。またプリズム装着による抑圧は予め連合野の一部を破壊しておくと生じなかった。ちなみに単眼遮蔽による抑圧は、この部位を予め破壊しておいても普通に誘発された。また神経系に特異的なある種の細胞接着因子を欠損するマウスでは、プリズム装着による抑圧は阻害されるのに対し、単眼遮蔽による抑圧は誘発された。これらの結果は、プリズム装着による視覚野抑圧は、これまで全く報告されていない新しいタイプの経験依存的可塑性であり、ヒゲ入力と視覚入力を統合する高次感覚連合野機能の働きに依存することを示している。
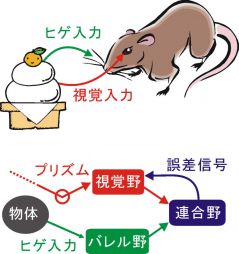
図3 連合野における視覚情報とヒゲ情報の統合
マウスは外界を認識するのに視覚とヒゲ情報を用いる。プリズム眼鏡により視覚のブレを起こすと両者の食い違いが生じ、統合が阻害される。このとき視覚野応答が抑圧される。この抑圧は、単眼遮蔽による抑圧と異なり、ヒゲを予め切っておいたり連合野を破壊しておくと消失する。
体性感覚野では末梢神経の一部が損傷されると、対応する体性感覚野脳地図の改変が生じたり、触覚が痛みに質的に変化する現象(神経因性疼痛)が生じることが知られている。これらは、皮膚に加わる大きな物理的な外力を回避するための生体防御機構として捉えることができる。この代償的可塑性は神経切断後1時間以内という速さで生ずるが、中枢が末梢神経の切断をどのようにして検出し、素早く可塑性を生じさせるのかはよく判っていない。我々は経頭蓋フラビン蛋白蛍光イメージングによって精度よく体性感覚野活動が記録できることの利点を生かし、体性感覚野の神経切断後の代償的可塑性を再現することに成功した。さらにこの実験系を用いて代償的可塑性のトリガーメカニズムを解析した。その結果、末梢神経切断によって極低頻度の自発発火が中枢に伝わらなくなること、また内因性GDNFが末梢神経の極低頻度発火を制御していることを見出した。さらに神経切断後数時間という速さで、触覚性の感覚信号が痛覚性の感覚信号へと質的な変化を起こすことを見出した(図4)。実は体性感覚野の応答を観察するだけでは、それが触覚によって生ずるのか痛覚によって生ずるのかは区別できないが、触覚は刺激と同側の脊髄後索を通って体性感覚野に到達するのに対し、痛覚は刺激と対側の脊髄視床路を通る。従って、これらの経路の部分破壊と体性感覚野のイメージングを組み合わせると、感覚情報が触覚から痛覚に変化したことが証明できる。またこのような感覚情報の質的な変化は脊髄レベルで生ずるので、様々な薬剤を投与することによって、そのメカニズムを追及することが可能である。
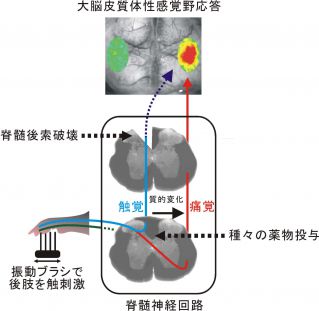
図4 神経因性疼痛の始まり
皮膚からの感覚神経の一部が障害されると周辺の感覚神経の感度が高まる。このとき、脊髄において、触覚から痛覚への感覚情報の質的変換が生ずる。
ハーモニーの快い感覚は、振動数比が単純な複数の音からなる和音を聴かせた時の「心」の領域の現象であり、そのままでは脳科学の対象とすることができない。しかし和音は、各成分の周波数の最大公約数に対応する基本周波数に対する知覚を生じさせ、この基本周波数に対応する音が和音に含まれない場合でも基本周波数に対応する聴覚野の応答として記録可能である。我々はこの現象のメカニズムとして経験依存的可塑性があると考えている。即ち動物の鳴き声などの自然界に存在する音の多くは基本周波数とその倍音からなる。鳴き声などの自然音が動物に繰り返し提示されるときには基本周波数に相当する音も倍音と同時に提示される。それぞれの周波数に良く応ずる皮質ニューロン群は同時に発火する確率が高いので、これらのニューロン間の皮質内結合がヘッブの法則によって形成・強化される。従って基本周波数成分を持たない和音が提示されたときにも、この皮質内結合を介して基本周波数応答が生じたと考えられる。つまり、倍音だけを聴いても存在しないはずの基本周波数応答が出現するのは、何度も倍音と基本周波数の組み合わせを聴いたことがあるという経験によって生じるのである。このようなメカニズムは、マウスの鳴き声を確実に検出するために役に立つと考えられる。鳴き声を用いた原始的なコミュニケーションシステムは動物において特定の感情を呼び起こすことが知られている。我々は特定の和音を聴くと何らかの情感を呼びさまされるが、それはこの原始的なコミュニケーションシステムの痕跡が我々の音楽を楽しむ脳機能に残されているためかも知れない。
IV.今後の可能性
これまで経頭蓋フラビン蛋白蛍光イメージングとはどのようなものか、これを用いてどのような問題が解決できるかを説明してきた。さらに、この方法と他の研究手法とを組み合わせることによって、広範囲の問題に応用することができる。例えば2光子顕微鏡を用いた単一ニューロン活動の解析と組み合わせることによって神経回路網レベルの様々な知見を解析することができる。脳活動は記録するだけでなく、抑圧したり刺激したりすることによってより強固な結論を得ることができる。経頭蓋的に脳活動を抑圧するのはフラビン蛋白の退色を利用した経頭蓋光不活化法が使用できる。一方経頭蓋的に脳活動を引き起こすには、オプトジェネティクスを利用する方法もあるが、より簡単には経頭蓋電気刺激法を使用することができる。即ち一般に頭蓋骨の上から電流を流し込んだ場合、電気抵抗が低い脳脊髄液に電流が拡散し、局所的な刺激が不可能であるが、予め頭蓋骨を薄く削り、局所的な圧力に応じて頭蓋骨が変形するようにしておけば、電極を押し当てた部位のクモ膜下腔を一時的に潰すことができる。このとき電極と頭蓋骨を介して流れ込んだ電流は脳脊髄液に拡散せずに脳実質に流れ込むので経頭蓋電気刺激が可能となる。さらに反応特性の差を利用した差分イメージング法、学習実験、脳切片における経験依存的可塑性の解析、覚醒マウスにおける脳活動の解析などを組み合わせ、経頭蓋フラビン蛋白蛍光イメージングをより広範な脳科学の問題に応用することができるはずである。
